

设计体验师Weifeng,Ben🧧
罗伊氏乳酸杆菌作为有前景的抗病毒药物的作用和潜在机制
31 丨5个月前
罗伊氏乳酸杆菌具有多种有益作用,包括维持体内平衡和肠道微生物的平衡、激活免疫系统和调节免疫反应。由于益生菌的有益作用,各种各样的益生菌已被开发为动物和人类健康的益生菌剂。病毒性疾病每年给牲畜造成严重的经济损失,对动物来说仍然是一个巨大的挑战。此外,预防和控制病毒性疾病的策略有限。病毒通过皮肤和粘膜表面进入宿主,其中有数亿微生物定植。罗伊氏乳酸杆菌的抗病毒作用已被证实,包括通过各种益生菌、益生菌代谢物和宿主信号通路调节化学、微生物、物理和免疫屏障。阐明益生菌的抗病毒机制意义重大,但还远远不够。本综述的主要目的是讨论益生菌的抗病毒作用和潜在机制,并为开发新型抗病毒药物提供目标。
人类和动物体内寄生着数以亿计的微生物,远远超过宿主细胞的数量,有人认为微生物的数量与宿主细胞相当(Egert and Simmering,2016)。这些微生物绝大多数共同栖息在胃肠道内(Sender et al., 2016 ; Heintz-Buschart and Wilmes, 2018 ; Mishra et al., 2021)。益生菌对于维持肠道微生物的平衡至关重要。粮食及农业组织 (FAO) 和世界卫生组织 (WHO) 将益生菌定义为:益生菌是活的微生物,当以足够量服用时,可给宿主带来健康益处(WHO/FAO, 2002 ; Drago et al., 2010; Kahouli et al., 2013)。益生菌因其有益作用而闻名,主要包括维持肠道微生物平衡、激活免疫系统和调节免疫反应(Hao et al., 2015 ; Gasbarrini et al., 2016 ; Kim et al., 2019)。由于其有益作用,近年来多种益生菌被广泛用于促进动物和人类健康(Kim et al., 2019 ; Teame et al., 2020 ; Rad et al., 2021)。已经评估了几种益生菌及其作为益生菌剂的效果。乳酸杆菌、双歧杆菌和酵母被广泛使用(Al-Ghazzewi and Tester, 2016 ; Wieers et al., 2020 ; Di Pierro and Pane, 2021 ; Giannakou et al., 2021)。
病毒性疾病仍然是人类和动物面临的巨大挑战(VanderWaal and Deen,2018)。尽管疫苗接种是预防病毒感染的最重要选择,但不断演变的流行病和现有疫苗之间的差异使得疫苗接种的效果降低。此外,目前尚无针对新出现或重新出现的病毒的疫苗。病毒通过皮肤和粘膜表面进入宿主,大量微生物在此定植(Schmidt 等人,2018 年;Lunjani 等人,2019 年;Xu 和 Li,2019 年;Peixoto 等人,2021 年)。益生菌对宿主抑制传入病原体感染至关重要(Lloyd-Price 等人,2016 年;Piewngam 等人,2018 年)。大量文献表明罗伊氏乳酸杆菌具有抗病毒作用(Pradhan 等,2021;Salaris 等,2021)。在本综述中,我们总结并讨论了益生菌抗病毒作用的机制,为抗病毒策略的使用开辟了新的视角,为抗病毒药物的研究和开发提供了新的靶点,并有助于为基于益生菌的抗病毒药物提供更清晰的背景。
益生菌有助于形成化学屏障,发挥抗病毒作用
化学屏障是病原微生物侵入肠道后遇到的第一道屏障。病原体必须突破化学屏障才能进入上皮。益生菌利用宿主中的碳水化合物产生各种代谢物,包括抗菌肽 (AMPs)、过氧化氢 (H 2 O 2 )、乳酸、短链脂肪酸 (SCFAs) 和细胞外囊泡 (EV)(Knezevic et al., 2005;Gosmann et al., 2017;Vieira et al., 2017;Tyssen et al., 2018;Nahui et al., 2019)。研究表明,益生菌释放的代谢物具有抑制病毒感染的重要保护活性。这些代谢物形成不利于病毒繁殖的微环境(图 1A)。
益生菌与病毒之间的相互作用。(A)益生菌通过构成或调节化学、微生物、物理和免疫屏障对病毒感染发挥保护作用。益生菌可以产生抗病毒代谢物,通过与病毒结合或竞争进入受体来阻止病毒入侵,调节 IEC 的紧密连接,并调节粘膜免疫反应。(B)益生菌调节全身免疫反应。
AMP 是一种短的、带正电荷的两亲性肽,具有广泛的抗菌活性,可对抗细菌、病毒、真菌和原生动物(Boparai 和 Sharma,2020 年;Bosch 和 Zasloff,2021 年)。近年来,AMP 因其治疗潜力而备受关注。细菌素是从细菌中提取的 AMP(Fry,2018 年)。虽然细菌素的抗菌作用已被某种程度上揭示,但其抗病毒作用仍有待进一步研究。肠球菌产生的肠素 B被证明能抑制流感病毒(H1N1 和 H3N2)在 Madin-Darby 犬肾细胞中的细胞病变作用(Ermolenko 等人,2019 年)。据推测,细菌素可以导致病毒颗粒聚集,通过与宿主细胞受体结合来阻断病毒颗粒或抑制病毒复制周期中的关键步骤(Wachsman et al., 2003)。细菌素的结构多样,但抗病毒机制相似。
在阴道黏膜中起保护作用的益生菌以罗伊氏乳酸杆菌属为主。大量研究表明,罗伊氏乳酸杆菌释放的代谢产物对人类免疫缺陷病毒(HIV)有抗病毒作用。罗伊氏乳酸杆菌产生的代谢产物之一H 2 O 2对许多病毒都有毒性,包括HIV。有报道称,HIV阳性女性中产H 2 O 2的乳酸杆菌的流行率低于HIV阴性女性(Knezevic et al.,2005)。另有研究表明,H 2 O 2能抑制CEM(一种人类T淋巴母细胞系)细胞中的HIV复制(Klebanoff and Coombs,1991)。健康女性阴道中存在的产H 2 O 2的益生菌被认为是维持健康的一些细菌。罗伊氏乳酸杆菌对HIV的破坏是由于H 2 O 2的毒性所致。
以往关于罗伊氏乳酸杆菌类益生菌抗病毒作用的研究主要集中在H 2 O 2上,但最近的研究表明,乳酸是阴道黏膜中罗伊氏乳酸杆菌产生的主要抗病毒因子( O'Hanlon等,2011;Liu等,2021)。乳酸是碳水化合物的最终产物,是乳酸杆菌的重要代谢产物,具有抗HIV活性(Gosmann等,2017;Nahui等,2017;Tyssen等,2018)。产乳酸菌抗HIV作用的机制可能是多因素的。首先,乳酸可能与病毒结合并影响病毒的功能,从而限制病毒侵入宿主细胞。其次,乳酸可以破坏病毒包膜,裂解包膜病毒的病毒体(Tachedjian等,2017)。除HIV外,乳酸对单纯疱疹病毒(HSV)也有明显的抑制作用(Conti等,2009)。已证实乳酸能成功干扰细胞中的病毒复制,并且杀病毒活性和复制抑制都与酸性pH值相关。
结肠中的微生物利用膳食碳水化合物作为底物产生 SCFA,最终产物主要是乙酸盐、丙酸盐和丁酸盐(Wong 等人,2006 年)。乳酸杆菌和双歧杆菌均具有产生 SCFA 的能力( LeBlanc 等人,2017 年)。据报道,罗伊氏乳酸杆菌可调节丁酸盐代谢,从而增强宿主对流感病毒感染的抵抗力。相关性分析显示,丁酸盐与病毒载量呈负相关(Lu 等人,2021 年)。罗伊氏乳酸杆菌产生的 SCFA 也是肠道上皮增殖的潜在调节效应物(Wang 等人,2018 年)。此外,SCFA 不仅可以作用于肠道,还可以作为通讯化学信号通过血液循环运输到远处部位。例如,在感染呼吸道合胞病毒 (RSV) 的小鼠中,施用益生菌混合物后,产生 SCFA 的细菌数量和血清 SCFA 水平增加 ( LeBlanc 等人,2017 )。同样,共生微生物群诱导的抗病毒反应也报告了类似的结果 ( Antunes 等人,2019 )。
乳酸杆菌可以和其他细菌和哺乳动物细胞一起产生胞外载体(EVs),这对于细菌和细胞之间的通讯至关重要。已证明L. crispatusBC3 和L. gasseri BC12 通过产生 EVs 保护人宫颈阴道和扁桃体组织免受 HIV-1 感染( Nahui et al., 2019)。EVs 通过减少 HIV-1 进入/附着于靶细胞来抑制 HIV-1 感染。此外,罗伊氏乳酸杆菌可以通过促进杯状细胞分泌黏蛋白在肠上皮上方形成黏蛋白层来改善屏障功能,保护黏膜免受病毒附着(Engevik et al., 2019 ; Lorella et al., 2020)。例如罗伊氏乳酸杆菌混合物VSL#3诱导结肠上皮细胞表达黏蛋白,并促使杯状细胞分泌黏蛋白阻止病原体附着(Caballero-Franco et al.,2007)。相关研究证实,罗伊氏乳酸杆菌可以分泌胞外蛋白,削弱病原体的粘附,保护肠道细胞(Liu et al.,2020)。病毒是细胞内病原体,需要宿主机制才能复制,这些代谢产物通过阻止病毒附着在宿主细胞上或直接杀死病毒而发挥抗病毒作用,是一种广谱抗病毒机制。综上所述,罗伊氏乳酸杆菌通过产生抗病毒代谢产物或刺激杯状细胞产生黏蛋白,增强化学屏障,维持宿主健康。
罗伊氏乳酸杆菌构成微生物屏障,发挥抗病毒作用
罗伊氏乳酸杆菌除了产生一些抗病毒代谢产物外,还可以通过与病毒相互作用或竞争细胞受体来直接抑制病毒,从而抑制病毒进入宿主细胞(图 1A)(Lievin-Le and Servin,2014)。
多项研究表明,罗伊氏乳酸杆菌可以在体外抑制水泡性口炎病毒(VSV)的感染。有研究表明,将罗伊氏乳酸杆菌预先孵育细胞单层或将罗伊氏乳酸杆菌与VSV一起孵育均可降低VSV滴度(Botic等,2007)。罗伊氏乳酸杆菌抑制VSV复制的一个可能机制是罗伊氏乳酸杆菌与病毒竞争细胞结合,干扰病毒附着或进入;另一个可能的机制是罗伊氏乳酸杆菌可以特异性或非特异性地捕获VSV。以上是罗伊氏乳酸杆菌抑制VSV感染的两种可能机制,更多的机制尚待阐明。枯草芽孢杆菌是具有良好抗病毒和免疫调节特性的益生菌之一。本研究证明枯草芽孢杆菌OKB105通过竞争进入受体抑制猪传染性胃肠炎病毒(TGEV)进入猪肠道上皮细胞系(IPEC-J2)(Wang et al.,2013)。另一项研究表明屎肠球菌NCIMB 10415通过益生菌与猪流感病毒(SIV)的直接相互作用有效抑制SIV感染(Wang et al.,2013)。已发表的文献分析表明罗伊氏乳酸杆菌的抗病毒作用具有菌株依赖性,此外罗伊氏乳酸杆菌还能限制病原体对营养资源的获取。此外,有研究表明无菌小鼠由于粘膜表面缺乏微生物定植,其粘膜免疫系统尚未成熟(Shen et al.,2014)。微生物的定植也促进宿主黏膜免疫系统的成熟,罗伊氏乳酸杆菌构成的微生物屏障对抑制病毒感染至关重要,为未来新型抗病毒策略的开发提供了思路。
罗伊氏乳酸杆菌增强物理屏障,发挥抗病毒作用
物理屏障由表皮和粘膜组成(Karst,2016)。罗伊氏乳酸杆菌通过增强肠上皮细胞(IEC)之间的紧密连接(TJ),形成抵御病毒入侵的防御屏障(图1A)。
TJ 是细胞间连接的重要形式,也是粘膜屏障的最重要结构(Otani and Furuse,2020 年)。IEC 通过 TJ 蛋白相互连接,TJ 蛋白是在上皮细胞和内皮细胞之间形成的具有防御功能的膜蛋白复合物(Gunzel and Fromm,2012 年)。据报道,不同种类的益生菌通过调节 TJ 蛋白的产生来保护宿主免受病毒感染(Kammani and Kim,2019 年;Lorella 等,2020 年)。Lactiplantibacillus ( Lp . ) plantarum、Weissella cibaria或Latilactobacillus ( Ll. ) sakei可以通过维持 TJ 蛋白的活性来保护 IEC 免受病毒感染。表皮细胞之间的 TJ 蛋白对于维持粘膜屏障的完整性是必不可少的。完整的黏膜屏障可以阻止病原体或病原体的抗原成分进入肠道固有层。TJ 蛋白缺乏会引起上皮屏障破坏,导致微生物易位至黏膜表面,破坏微生物稳态。此外,已证实罗伊氏乳酸杆菌( Bf . )治疗可促进小鼠小肠上皮增殖 ( Ishizuka et al., 2009)。小肠上皮细胞的增殖促进了新细胞的产生和坏死细胞的脱落。上皮细胞形成的物理屏障不断自我更新,增强了黏膜物理屏障抵抗病原体的能力。此外,罗伊氏乳酸杆菌还能调节跨上皮电阻和上皮通透性 ( Krishnan et al., 2016 )。肠道微生物环境的平衡是人类和动物维持体内平衡的必要条件,综上所述,罗伊氏乳酸杆菌可以通过维持肠道通透性和黏膜屏障完整性来抑制病毒感染。
罗伊氏乳酸杆菌调节免疫屏障发挥抗病毒作用
罗伊氏乳酸杆菌最有益的作用之一是调节免疫反应(Shida 和 Nanno,2008 年;Azad 等,2018 年)。免疫屏障是宿主抑制病毒感染的最后一道防线,由粘膜和全身免疫反应组成(图 1A、B)(Ashraf 和 Shah,2014 年;La Fata 等,2018 年)。
粘膜免疫反应
一般来说,胃肠道中的淋巴组织和免疫细胞数量比身体其他部位的总和要多。粘膜免疫系统(MIS)又称粘膜相关淋巴组织(MALT),主要由免疫组织、细胞和分子组成。粘膜与外界环境直接联系,参与复杂的免疫反应(Pelaseyed et al.,2014)。各种免疫细胞参与先天免疫反应。自然杀伤(NK)细胞是先天免疫的关键成员,在识别和杀死靶细胞、调节免疫反应中起关键作用。NK细胞的激活和增殖可以有效限制病毒复制。据报道,Lp. plantarum 06CC2 可降低小鼠脑中 HSV-1 病毒产量(Matsusaki et al.,2016)。Lp . plantarum 06CC2 通过增加派尔淋巴结 (PPs) 中 IL-12 和 IFN-γ 的 mRNA 表达来增强免疫调节活性。IL-12 和 IFN-γ 可以有效激活 NK 细胞和巨噬细胞。巨噬细胞和 NK 细胞相互协作清除病毒。
模式识别受体(PRR)在微生物与宿主之间具有广泛的相互作用。PRR是宿主细胞识别病原体的特化分子。目前,PRR主要包括Toll样受体(TLR)、核苷酸结合寡聚化结构域(NOD)样受体(NLR)、视黄酸诱导样受体(RLR)、C型凝集素受体(CLR)、黑色素瘤2型缺失(AIM2)样受体(ALR)和环鸟苷酸-腺苷酸合酶(CGA)。TLR成员可识别多种病原体,如细菌、病毒和真菌。已有研究证明,鼠李糖乳杆菌GG(LGG)通过TLR3信号通路对轮状病毒(RV)感染引起的腹泻具有治疗作用(Aoki-Yoshida et al.,2016)。罗伊氏乳酸杆菌也通过激活TLR3信号通路表现出抗病毒作用(Indo et al., 2021)。TLR3主要识别内体中的核酸,TLR3的激活会增加I型IFN的表达。罗伊氏乳酸杆菌除了激活TLR3外,还能激活其他PRR,例如嗜酸乳杆菌可通过诱导小鼠树突状细胞(DC)中病毒免疫防御基因的表达增强抗病毒作用(Weiss et al., 2010),病毒诱导的免疫防御基因的表达依赖于TLR2通路的激活。此外,植物乳酸杆菌可以通过NLR(NOD2)和TLR(TLR2)通路抑制小鼠肺病毒(PMV)感染( Rice et al., 2016)。 NOD2 和 TLR2 均介导重要的先天免疫反应来抑制病毒感染。多项研究表明,罗伊氏乳酸杆菌混合物也具有免疫调节活性。罗伊氏乳酸杆菌混合物对下调促炎细胞因子(如 IL-6、IL-8 和 IL-1β)的表达有重要影响(Macpherson 等人,2014 年)。主要影响包括上调 TLR3、丝裂原活化蛋白激酶和因子 κB (NF-κB) 的表达。此外,Lp. plantarum、W. cibaria和Ll。sakei可通过激活 TLR 和 NF-κB 通路来调节 IEC 中 poly(I:C) 诱导的先天抗病毒免疫反应 ( Kanmani and Kim, 2019 )。NF-κB 信号通路与促炎细胞因子的产生有关。在病毒感染期间,激活 NF-κB 信号通路可增加促炎细胞因子的产生并调节免疫细胞的功能。
干扰素是抗病毒免疫和调节免疫系统稳态的重要介质(Stefan et al.,2020)。据报道,罗伊氏乳酸杆菌通过诱导肠道抗原呈递细胞(APC)产生I型干扰素来抑制病毒感染。此外,罗伊氏乳酸杆菌已被证明可通过在早期诱导大量IFN-β,在晚期激活JAK-STAT1通路对IPEC-J2细胞发挥抗TGEV作用,而激活的JAK-STAT1通路会增加一些抗病毒蛋白的转录和表达。
病毒的有效清除依赖于先天性和适应性免疫反应的协调。越来越多的证据表明,罗伊氏乳酸杆菌可以通过刺激适应性免疫反应保护宿主免受病毒感染。罗伊氏乳酸杆菌显著降低粪便、回肠和肠系膜淋巴结 (MLN) 中的 PCV2 病毒载量,并增加回肠中的免疫球蛋白 A (IgA)(Karaffova 等,2017)。IgA 是粘膜免疫系统的主要成分,广泛分布于宿主粘膜组织的粘膜分泌物中,可抑制病毒附着在上皮细胞上,减缓病毒复制,在免疫屏障中发挥重要作用。越来越多的证据表明,不仅活的罗伊氏乳酸杆菌而且热灭活的罗伊氏乳酸杆菌都具有免疫调节作用。例如罗伊氏乳酸杆菌通过鼻腔内给药诱导病毒特异性抗体,对 IFV 感染具有广泛的保护作用 ( Jung et al., 2017 )。分泌型 IgA 可以特异性地与病毒结合,抗体捕获的病毒可以被吞噬细胞摧毁。
全身免疫反应
定植于黏膜表面的罗伊氏乳酸杆菌也具有针对病毒感染的远端保护活性(图1B)。已证实罗伊氏乳酸杆菌通过调节局部黏膜免疫具有抗病毒作用。最近的一项研究表明,罗伊氏乳酸杆菌还可以通过调节全身免疫反应来保护小鼠免受病毒感染(Mudronova等,2018)。此外,口服Lc. rhamnosusLA68可诱导脾脏或血液中一些细胞因子的表达,激活Th1型免疫反应(Dimitrijevic等,2014)。罗伊氏乳酸杆菌或其代谢产物可以激活免疫细胞移动远端位置来介导抗病毒反应,或通过刺激免疫细胞释放细胞因子通过血液和淋巴循环作用于远端。
罗伊氏乳酸杆菌是肠道微生物的主要成员之一,具有免疫调节作用。罗伊氏乳酸杆菌鼻腔接种IFV后,可改善小鼠临床症状,降低死亡率。罗伊氏乳酸杆菌可增强肺和脾脏中的NK细胞活性,并增加细胞因子(IFN-γ、IL-2、IL-12和IL-18)的表达(Kawahara等,2015)。粪肠球菌CECT7121可诱导树突状细胞强烈活化,并分泌高水平炎症细胞因子(IL-12、IL-6、TNF-α和IL-10)(Molina等,2015)。这些细胞因子对抗病毒先天免疫反应至关重要。IL-10是一种已知的炎症和免疫抑制因子,可以调节免疫反应。由于病毒感染,IL-10表达增加可减轻炎症对宿主的损害。树突状细胞(DC)是免疫系统中的哨兵细胞,起着APC的作用。它们也是先天免疫和适应性免疫之间的桥梁,可以直接激活T细胞和B细胞或迁移到肠系膜淋巴结。此外,口服罗伊氏乳酸杆菌可通过在肺中产生高水平的IL-12和IFN-γ赋予IFV保护活性。IL-12是先天免疫和适应性免疫的关键调节分子。
干扰素是先天免疫的介质,在抵抗病毒感染中起关键作用。据报道,罗伊氏乳酸杆菌产生的双链 RNA 可触发树突状细胞 (DC) 产生 IFN-β ( Kawashima 等人,2018 )。罗伊氏乳酸杆菌通过增加 IFN-α 和 ISG 的表达来调节人体对 IFV 的免疫反应 ( Sugimura 等人,2015 )。此外,口服罗伊氏乳酸杆菌对小鼠的 H1N1 流感病毒感染具有保护作用 ( Maeda 等人,2009 )。罗伊氏乳酸杆菌的有益作用是通过诱导宿主细胞产生 I 型干扰素来介导的。罗伊氏乳酸杆菌还可以通过刺激 AM 产生 IFN 来保护宿主免受 RSV 感染 ( Ji 等人,2021 )。 IFN信号通路的激活会增加ISG的表达,ISG是IFN的主要抗病毒效应物。越来越多的研究表明,ISG靶向病毒复制的不同阶段来抑制病毒感染。据报道,罗伊氏乳酸杆菌可以通过上调Mx1和信号转导和转录激活因子1(STAT1)基因显著降低鸽子粪便和脾脏中的鸽圆环病毒(PiCV)病毒载量(Tsai等人,2021年)。ISG之一的粘液病毒抗性1(Mx1)可以在病毒侵入细胞后阻断其核酸的早期转录(Zurcher等人,1992年)。口服罗伊氏乳酸杆菌可以通过产生I型IFN和ISG(包括IFNAR1、Mx2、OAS1、OAS2、RNase L和IFITM3)来提高对RSV感染的抵抗力(Garcia-Castillo et al., 2020)。这些分子增强了抑制小鼠RSV感染的能力。罗伊氏乳酸杆菌还通过上调IFN和ISG的表达具有预防RSV感染的潜力(Eguchi et al., 2019 )。此外,还鉴定出一个具有RSV抗病毒活性的罗伊氏乳酸杆菌候选蛋白SRCAP 。但SRCAP蛋白抑制RSV复制的具体功能有待进一步确定。此外,罗伊氏乳酸杆菌 还可以通过增加Mx1和Oas1a基因的表达来保护小鼠免受 IFV 感染,这两个基因对于降低肺部病毒滴度至关重要(Nakayama 等人,2014 年)。上调 ISG 可有效抑制 IFV 感染。
此外,罗伊氏乳酸杆菌保护小鼠可抑制流感病毒和肠道病毒感染(Chen et al., 2017 )。粪肠球菌的保护作用与 MCP-1/CCR2 通路的激活有关,该通路可能是增强抗病毒免疫反应的关键介质。MCP-1 的表达水平与病毒载量呈负相关。
众所周知,有效消灭病毒依赖于适应性免疫。用罗伊氏乳酸杆菌治疗的小鼠产生了抗体IL-4、IL-12和IFN-γ,并免受H1N1流感病毒的攻击(Mahooti et al., 2019)。IL-4可诱导Th2免疫反应,而IFN-γ调节Th1免疫反应。Th1和Th2之间的平衡对于宿主免疫系统的稳态很重要。口服罗伊氏乳酸杆菌可在支气管肺泡灌洗液中诱导sIgA和中和抗体,并抑制肺部病毒增殖(Kawashima et al., 2011)。此外,口服罗伊氏乳酸杆菌可延长小鼠感染IFV后的存活时间,且sIgA和Th1类细胞因子(IL-2和IFN-γ)显著升高(Song等,2016 )。因此,小鼠对IFV感染的抵抗力归因于鼠李糖乳杆菌M21激活了细胞免疫反应。
结论与观点
当今世界正面临多种新型传染病,其中病毒性疾病尤为严重,目前WHO已指出,严重的冠状病毒病2019(COVID-19)是对人类的一次大流行性挑战。疫苗接种是抑制病毒感染的重要工具,但针对新型病毒的疫苗研发存在滞后,且一些抗病毒药物也存在一些不良反应,开发新型抗病毒策略迫在眉睫。益生菌在人类和动物健康中的作用是近年来令人关注的话题,罗伊氏乳酸杆菌已被证明具有抗病毒作用,代表着未来病毒性疾病的安全替代预防药物(Stavropoulou and Bezirtzoglou,2020;Sundararaman et al.,2020)。然而,目前对罗伊氏乳酸杆菌抗病毒作用的研究较少,临床数据仍不足。目前罗伊氏乳酸杆菌的抗病毒作用主要在体外或小鼠模型中进行,需要在病毒的天然宿主中进行验证。更多动物模型有待建立。越来越多的证据表明益生菌的抗病毒作用与给药途径有关。罗伊氏乳酸杆菌通常的给药途径是口服,油剂更容易服用,可滴到鼻腔或口腔内。但最近的研究表明,鼻腔内或舌下给药也能降低病毒载量,提高动物的存活率。鼻腔内和舌下途径可能是罗伊氏乳酸杆菌口服给药的替代方法。罗伊氏乳酸杆菌的剂量也会影响抗病毒作用。同一种罗伊氏乳酸杆菌菌株在不同剂量下可诱导不同的免疫反应。过量服用罗伊氏乳酸杆菌可能会增加免疫抑制的风险。因此,在使用罗伊氏乳酸杆菌类抗病毒药物时,应更加注意罗伊氏乳酸杆菌的种类、剂量和给药途径。此外,还应进行更多的临床研究,以揭示哪些罗伊氏乳酸杆菌或其组合对特定病毒最有效。
免疫系统的稳态有助于宿主抵抗病毒感染。已证明罗伊氏乳酸杆菌不仅可以激活局部免疫反应,还可以激活针对病毒感染的全身免疫反应。其确切机制仍未得到证实。虽然罗伊氏乳酸杆菌的抗病毒机制已被部分揭示,包括罗伊氏乳酸杆菌成分或代谢物以及相应的宿主 PRR 参与的抗病毒作用(如表 1所示),但有必要更详细地了解罗伊氏乳酸杆菌的抗病毒作用,并需要进行更广泛和准确的研究,以使用新的分子工具和技术阐明罗伊氏乳酸杆菌抗病毒作用的潜在机制。了解罗伊氏乳酸杆菌背景下人类和动物的抗病毒免疫有利于制定新的抗病毒策略。罗伊氏乳酸杆菌的抗病毒作用将成为未来研究的一个有趣领域。罗伊氏乳酸杆菌,德国直邮,品质保证。
杜塞尔多夫
 0
0
 1
1
 93
93  广告
广告
汉语教学
9
 广告
广告
市中心诚聘足疗按摩师
6
 广告
广告
OCCIDENT线上保险办理
31
0
 广告
广告
【顺通通渠】 电脑检测 免费评估
31
 广告
广告
🔥🔥国际学校车程10分钟内,景观无敌的半山海景别墅
23
9-10亿韩币
 广告
广告
房屋出售
29
0
 广告
广告
小巴黎 11区日餐自助请鱼生师傅 会讲国语 备本人拘留
6
 广告
广告
税务优化:比较出租房空租及带家具出租
77
 广告
广告
加拿大冰酒,红酒对华贸易招聘兼职销售!
6
 广告
广告
三岁半纯种蓝猫公猫找新家
18
200
 广告
广告
专业验房师
31
 广告
广告
留学生回国转租❗️600欧全包米兰单间7月20日起租到10月底 ❗️正规合同‼
21
650
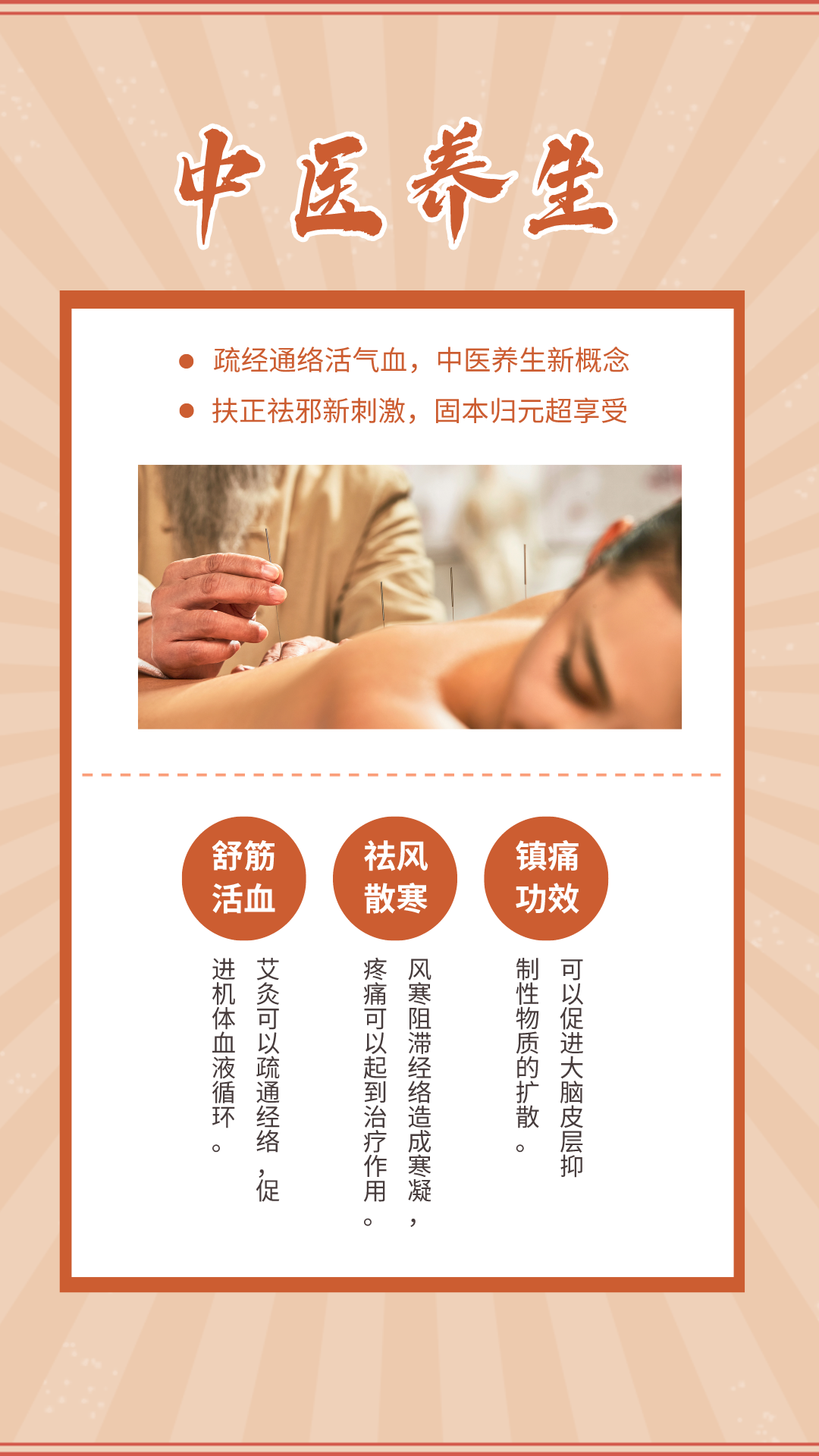 广告
广告
NARIS爱丽舍医疗美容会所
31
120~不等
 广告
广告
澳大利亚黄金树西拉
37
156
 广告
广告
Pantai Malindo Half Marathon 2025
17
0
 广告
广告
#全球房产eXpRealtor蔡凯傑BentonTsai
26
 广告
广告
Mallorca中西翻译、陪同、代办
31
 广告
广告
华会社信息平台 收费标准(2023)
31
 广告
广告
出租办公室及专业用房 191 平方米 巴黎 2区 (75002)
24
6.330 欧元
 广告
广告
圣彼得堡|魔饭大叔(加工外卖)
31
 广告
广告
野生干冻羊肚菌
36
200$
 广告
广告
台灣昆昉公司招聘資訊
6
15000000
 广告
广告
面包蜜语招聘
6
 广告
广告
海华233墨尔本站广告位招商! 海华233墨尔本运营商 !
31
 广告
广告
高档化妆品销售顾问,综合薪水3000-4000 新
6
4000
 广告
广告
超值三层公寓式住宅,(可以部分用作医生等自由职业者办公室)海景,游泳池,网球场,停车场
23
595 000 €
 广告
广告
【普罗旺斯风情豪宅】7室6卧,4700㎡超大花园,带泳池和按摩浴缸!
23
750 000€
 广告
广告
近市区合理房价出租
29
490
 广告
广告
🇹🇭芭提雅华人汽修🇨🇳RMY Garage Pattaya
31
1,000
 广告
广告
🇺🇸美国太阳能公司 Palmetto Solar 华人团队
30


